Mitogen-activated Protein Kinases in the Development of Normal and Diseased Kidneys
Article information
Abstract
Mitogen-activated protein kinases (MAPKs) play important roles in various cellular functions including proliferation, differentiation, and apoptosis. We showed that MAPKs are developmentally regulated in the rat kidney. p38 MAPK (p38) and extracellular signal-regulated kinase (ERK) were strongly expressed in the fetal kidney, whereas c-Jun N-terminal kinase (JNK) was detected predominantly in the adult kidney. The inhibition of p38 or ERK in organ culture resulted in reduced nephron formation with or without reduced kidney size. On the other hand, persistent fetal expression pattern of MAPKs, i.e., upregulation of p38 and ERK and downregulation of JNK, was observed in the cyst epithelium of human renal dysplasia, ovine fetal obstructive uropathy, and pcy mice, a model of polycystic kidney disease. Furthermore, activated p38 and ERK induced by cyclic stretch mediated proliferation and TGF-β1 expression in ureteric bud cells, probably leading to cyst formation and dysplastic changes. Inhibition of ERK slowed the disease progression in pcy mice. Finally, ERK and p38 were inactivated in the early embryonic kidney subjected to maternal nutrient restriction, characterized by reduced ureteric branching and nephron number. Thus, MAPKs mediate the development of normal and diseased kidney. Their modulation may result in novel therapeutic strategies against developmental abnormalities of the kidney.
Introduction
The mitogen-activated protein kinase (MAPK) family is serine/threonine kinases that comprise three major subgroups, i.e. extracellular signal-regulated protein kinase (ERK), p38 MAPK (p38), and c-Jun N-terminal kinases (JNK) [1]. Classically, ERK is thought to be activated by growth factors and vasoactive substances and to stimulate proliferation and differentiation. On the other hand, p38 and JNK are activated by cytokines and cellular stresses such as ultraviolet light, heat shock, hyperosmolality, etc. and have been implicated in growth inhibition and apoptosis [2]. It has become apparent, however, that their functions are not so specific and there are overlaps among the subtypes. MAPKs have transcription factors as their targets, and thereby regulate the induction of genes important in the biological response of the cell. They also regulate cytoplasmic targets such as phospholipase A2. MAPKs are activated by dual phosphorylation on tyrosine and threonine residues. MAPKs are phosphorylated by MAPK kinases that, in turn, are activated by the MAPK kinase kinases forming a cascade (Fig. 1).
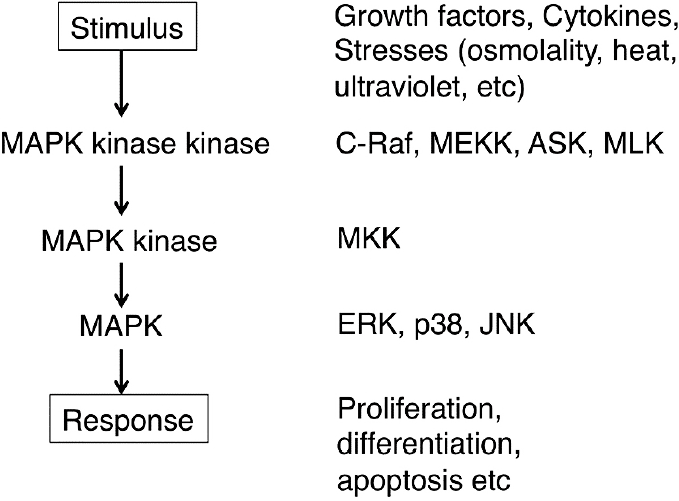
MAP kinase cascade. Mitogen-activated protein kinases (MAPKs), including ERK, p38, and JNK, are activated by upstream MAPK kinases (MAPKKs) by dual phosphorylation on tyrosine and threonine residues. MAPKKs are activated by MAPKK kinases (MAPKKKs), which are stimulated by various stimuli such as growth factors, stresses, etc.
While many studies have investigated the role of growth factors and genes during development of the kidney, not much had been known on the signaling pathways in the early 2000s. Since MAPKs mediate proliferation and apoptosis that are observed in a large scale in the developing kidney, we studied their expression and roles [3,4].
Expression in normal kidney
Both ERK and activated phosphorylated ERK (P-ERK) were present in the rat kidney throughout the stages [3]. The abundance of ERK was highest at embryonic stage and decreased gradually after birth (Fig. 2A). On the other hand, p38 and phosphorylated p38 (P-p38) were present almost exclusively in the embryonic kidney, with greatly diminished levels in early neonatal kidneys. They were undetectable in late neonatal and adult kidneys. The opposite pattern of expression was seen for JNK and phosphorylated JNK (P-JNK), which was strongly expressed in the adult kidney, was present with lower levels in neonatal kidneys but not detectable in the embryonic kidney.
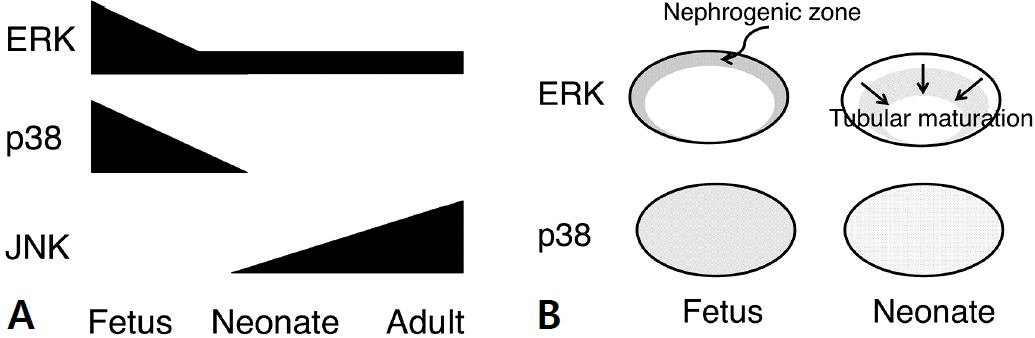
Expression of MAPKs during kidney development. (A) Temporal expression of MAPKs during kidney development. ERK and p38 are strongly expressed in the fetal kidney. ERK continues to be expressed throughout the stages with diminished levels. JNK is expressed predominantly in the adult kidney. (B) Spatial expression of MAPKs during kidney development. Expression of ERK correlates with kidney development, being strongly expressed in the nephrogenic zone during fetal stage and shifting to subnephrogenic zone to the medulla as tubules mature. p38 is diffusely expressed in the fetal kidney.
The spatial expression of ERK and P-ERK correlated with proliferation. Thus they were strongly expressed in the nephrogenic zone, where active ureteric branching and nephron formation were occurring (Fig. 2B). As the tubules mature, proliferation became more intense in subcortical region and the medulla. The localization of ERK and P-ERK also shifted in correlation to proliferation. p38 and P-p38, on the other hand, was observed diffusely in the embryonic kidney. Contrary to common belief, it did not correlate with the distribution of TUNEL-positive cells undergoing apoptosis, which was prominent in the nephrogenic zone. JNK was detected predominantly in the adult kidney localizing in podocytes and tubular cells. The expression of P-JNK was limited to distal tubules, collecting ducts, and podocytes.
Thus the expression of MAPKs in the rat kidney is unique during development. The similar spatial pattern was also observed in mouse kidney [5]. Although not all stages were examined, human and ovine kidneys seem to follow a similar pattern [6,7]. In summary, both the temporal and spatial expression of ERK coincides with the maturation of the kidney. The distribution of p38, on the other hand, is diffuse and correlates with proliferation rather than apoptosis. JNK is predominantly expressed in mature kidneys suggesting its role in the differentiation at a later stage or in the maintenance of the integrity of tubular epithelia.
Role of MAPKs during normal kidney development
We then investigated the role of ERK and p38 in an organ culture system [4]. Inhibition of MEK, an upstream kinase of ERK, by PD98059 attenuated nephron formation with minimal effect on kidney growth. On the other hand, both kidney growth and nephron formation were suppressed by a p38 inhibitor SB203580. Very few mesenchymal condensates were observed in kidneys incubated with SB203580. p38 inhibition also dramatically reduced proliferation and increased apoptosis compared with ERK inhibition, which explains the reduction in kidney size. The WT1 Wilms’ tumour suppressor gene expression is low in uninduced mesenchyme, and increases during mesenchyme-epithelium conversion in normal control metanephroi. In PD98 059-treated explants, WT1 expression was well observed. In SB203580-treated cultures, on the other hand, WT1-positive cells were distributed loosely in mesenchyme. Cells around ureteric buds, which show some signs of condensation, tended to have stronger WT1 expression.
While ureteric branching was not altered by the inhibition of ERK or p38 in our study, Fisher et al reported that a MEK inhibitor PD98059 inhibited ureteric branching [8]. The discrepancy may be explained by the fact that we cultured later stage of metanephroi compared with those in Fisher et al’s study. It is now known that ERK is a downstream signal of Ret, the receptor of GDNF that promotes ureteric branching. According to a recent study, genetic removal of ERK blocked branching while the ureteric epithelium continues to elongate [9]. Furthermore, enhanced epithelial adhesion following removal of ERK signaling sug gested that ERK promotes cellular motility through modulation of focal adhesion. p38 has also been shown to regulate branching morphogenesis lying downstream of integrin-linked kinase [10].
The role of JNK during kidney development was investigated by Osafune et al [11]. They showed that JNK lies downstream of WNT4, a secreted protein critical for nephrogenesis, and mediates mesenchymal-to-epithelial transformation of progenitor cells in mouse embryonic kid ney. As will be discussed later, JNK is also involved in the maintenance of progenitor cells mediated by BMP7 [12]. Thus although the expression level is low, JNK is important in the kidney development as well as ERK and p38.
MAPK in the development of abnormal kidney
Since MAPKs are necessary for kidney development, its dysregulation may lead to kidney malformation. We investigated the expression and role of MAPKs in human renal dysplasia, an ovine model of fetal obstructive uropathy, ureteric bud cell stretch, in vitro mimic of obstruction, pcy mice, a model of polycystic kidney disease (PKD), and kidneys of rat offspring exposed to maternal nutrient restriction.
1. Dysplasia
Renal dysplasia is one of the most common congenital anomalies of kidney and urinary tract. It is characterized by dysplastic tubules connected to cysts and surrounding undifferentiated tissue. Increased cell proliferation of dysplastic epithelia as well as prominent apoptosis in undifferentiated mesenchyme are the characteristic features. In normal human kidneys from 19 to 34-week fetuses, proliferation and apoptosis were prominent in the nephrogenic zone [6]. ERK and P-ERK were detected intensely in distal tubules and collecting ducts probably reflecting the advanced stage of development as speculated from the observation in rat kidney described above. Immature tubules and non-induced mesenchyme in the nephrogenic zone weakly expressed ERK. p38 and P-p38 were not detected in normal kidneys at any stage examined again probably because of relatively advanced developmental stage. In support of this, JNK and P-JNK were positive in tubular epithelia of normal fetal kidneys. A marker of proliferation PCNA was stained intensely in dysplastic tubules and weakly in cyst epithelia of dysplastic kidneys. Apoptosis was prominent in dysplastic tubules and cysts, and occasionally found in surrounding undifferentiated cells. Most strikingly, both p38 and P-p38 were intensely stained in dysplastic epithelia. p38 expression was also detected in areas of undifferentiated cells. Furthermore, contrasting to normal kidney, JNK and P-JNK were not detected in dysplastic epithelia. Dysplastic tubules and cysts expressed ERK and P-ERK. Thus p38 and ERK, mediators of proliferation, are upregulated and activated, while JNK is downregulated in dysplastic epithelia. Dysregulated MAPKs may cause hyperproliferation of dysplastic tubules leading to cyst formation.
2. Ovine fetal obstructive uropathy
Human renal dysplasia is frequently associated with urinary tract obstruction which is suggested to be one of the causative factors. In animals, prenatal obstruction of the urinary tract reproduces dysplastic changes similar to human dysplasia. In fetal lambs, obstruction at gestational ages 50-60 days produces a dysplastic kidney, whereas obstruction at more than 90 days of gestation generates hydronephrosis [13]. In adult animals, urinary tract obstruction leads to proliferation of tubule cells and interstitial fibrosis. The expression of P-ERK is reported to closely correlate with these events both temporally and spatially. A similar mechanism may be operative in fetal obstructive uropathy. We therefore examined the expression of MAPKs in ovine neonatal kidney obstructed at gestational age 60 days [7]. Cysts and undifferentiated mesenchyme and fibromuscular collars were observed along with increased expression of TGF-β and α-smooth muscle actin. Proliferation and apoptosis were prominent in tubules, cysts, and occasional interstitium. Similar to human dysplasia, ERK, P-ERK, and p38 was upregulated in cyst epithelia along with Pax2, a transcription factor known to be upregulated in cyst epithelia [14]. While JNK is expressed in tubules of normal neonatal kidney, it was not detected in tubules or cysts of obstructed kidney. TGF-β1 in cysts and interstitium correl ated with the expression of P-ERK. Overall, persistent fetal expression pattern of MAPKs was observed in cyst epithelium. P-ERK correlated both spatially and temporally with proliferation and TGF-β1 expression, suggesting that ERK may contribute to cyst formation as well as fibrosis in the fetal obstructive uropathy.
We next investigated the effect of vesico-amniotic shunting in fetal obstructive uropathy (unpublished observation). Three phenotypes were obtained by shunting; hydronephrosis, multicystic dysplastic kidney, and hypoplastic kidney [15]. In these kidneys, P-ERK was detected in only collecting ducts in the same way as normal kidneys, and cysts were negative for P-ERK or TGF-β1. These findings suggest that activation of ERK is mediated by mechanical stretch due to obstruction, after the resolution of which TGF-β1 is no more expressed even if the dilation of tubules remains.
3. Cyclic stretch of ureteric bud cells
To prove the above hypothesis, we investigated whether cyclic mechanical stretching of ureteric bud cells, a mimic of the hydrodynamic derangement after fetal urinary tract obstruction, reproduces events seen in vivo [16]. Cyclic stretch activated p38 and ERK, stimulated proliferation, and upregulated Pax2 and TGF-ß1 expression at 24 hours. Stretch-induced Pax2 and TGF-β1 expression was abolished by inhibitors of MEK and p38. Apoptosis was induced at later time points, 48 hours. The stretch-induced apoptosis was also abolished by ERK or p38 inhibition as well as by anti-TGF-β1 antibody.
To conclude, cyst formation and fibrosis seen in fetal obstructive uropathy may be due to the activation and upregulation of ERK and p38, which result from mechanical stretch of ureteric bud cells due to obstruction.
4. DBA/2pcy / pcy ( pcy ) mouse model of polycystic kidney disease
We next investigated the expression of MAPKs in a murine model of PKD another model characterized by multiple cysts [5]. Although the gene responsible for pcy mouse is a causative gene for nephronophthisis, the disease progression resemble for that of autosomal dominant PKD in humans [17]. In a similar manner to human dysplasia and ovine fetal obstructive uropathy, cyst epithelium of pcy mouse showed upregulation of ERK and p38 and downregulation of JNK. These changes were also associated with increased proliferation and apoptosis.
This in vivo model allowed us to examine the effect of ERK inhibition. Firstly we cultured the embryonic day-14 kidney from pcy mice in the presence or absence of MEK inhibitor U0126. The size and number of cysts were significantly reduced by ERK inhibition (unpublished observation). However, in treated kidneys, the width of the nephrogenic zone and the number of glomerular structures were reduced. Thus in a similar manner to rat embryonic kidney, ERK inhibition suppressed nephrogenesis in pcy kidney. To circumvent this problem, we next administered an orally active MEK inhibitor PD184352 into adult pcy mice [5]. Seven-week administration to 10 week-old pcy mice significantly reduced cyst formation. The expression of ERK and P-ERK was also decreased in PD184352-treated mice. This was associated with the reduction in blood pressure, kidney weight, serum creatinine level, and water intake and an increase in urine osmolality. PD184352 did not affect upregulation of p38 and P-p38 or downregulation of P-JNK in pcy mice. These results opened up new therapeutic approaches to PKD as well as cyst formation in fetal obstructive uropathy. While ERK inhibition affects the normal kidney development, targeted drug delivery to dysplastic or stretched epithelia may become possible and circumvent the problem in the future.
One may wonder whether p38 inhibition may also alleviate cyst formation in pcy mice. We cultured embryonic pcy kidney in the presence of SB203580 (unpublished observation). Unexpectedly, SB increased the size and the number of cysts. p38 inhibition has been reported to augment progression of remnant kidney model by activating ERK and enhancing proliferation [18]. A similar mechanism may be operative and explain our observation.
5. Kidneys from offspring of nutrient-restricted rat dams
While congenital anomaly of kidney and urinary tract is the largest cause of childhood chronic renal failure, recently recognized developmental origins of health and diseases (DOHaD) concept may even have a bigger impact on renal health. Thus disturbed intrauterine organogenesis is considered to affect the health and disease in the adulthood. In the kidney, low birth weight is associated with a risk for hypertension and kidney disease. While low nephron number is thought to be the major cause, its mechanism has not been fully elucidated. We therefore investigated the mechanism in a rat model of maternal nutrient restriction [19]. We found that ureteric branching, an important determinant of nephron number, as well as kidney size was decreased by maternal nutrient restriction. Activated forms of ERK and p38 along with other signaling molecules PI3K, Akt, and β catenin, important in kidney development, were also inactivated or downregulated in the early embryonic kidney. Decreased ERK signaling as well as GDNF in the fetal kidney exposed to maternal nutrient restriction was also reported by other investigators [20]. As discussed in the previous section, ERK and p38 are known to be necessary for ureteric branching as well as nephrogenesis [4,8,21]. Since GDNF activates ERK and p38, their reduced activity may be partly due to the decrease in GDNF. However, its receptor Ret is expressed only in the tips of ureteric buds. Hence there probably exist a more general mediator of maternal nutrient restriction which most likely affects progenitor cells.
Upstream signal of MAPKs during kidney development
Besides GDNF described above, many growth factors are known to activate ERK and p38 including BMPs, FGF, IGF, HGF, PDGF, TGF-β, EGF, activin, LIF, and others. Since BMP7 knockout mice show a similar phenotype to the kidney treated with inhibitors of p38 or MEK, we investigated the relationship between BMP7 and ERK and p38 in a metanephric mesenchymal cell line [22]. BMP7 is expressed in ureteric buds and cap mesenchyme of the fetal kidney, mediating branching morphogenesis and survival and priming of metanephric mesenchyme. While dose-dependent effects of BMP7 via p38 activation in collecting duct cells have been reported, studies in metanephric mesenchymal cells were lacking [23]. We found that BMP7 differentially activated ERK and p38 in metanephric mesenchymal cells (submitted). Thus P-ERK was increased with low-dose BMP7 (0.25 nM) whereas p38 was activated by intermediate-dose BMP7 (1 nM). Cell proliferation was stimulated by low- and intermediate-dose BMP7 but inhibited by high-dose BMP7 (10 nM). Low-dose BMP7-mediated proliferation was inhibited by inhibitors of MEK and p38. Expression of cadherin-11, an adhesion molecule known to promote cell migration and compaction, was upregulated by intermediate-dose BMP7, which was inhibited under coexistence of SB203580 and PD98059. In metanephroi cultured with siRNA for Cadherin-11, the number and thickness of cap mesenchyme was reduced. This is reminiscent of the metanephroi grown under SB20 3580 in our previous study [4] suggesting that the main target of p38 blockade was BMP7 pathway. In support of this, previous studies reported the involvement of JNK and p38 as the mediators of BMP7 in the maintenance of mesenchymal progenitor cells [12,24]. Our results demonstrate that ERK also plays a role in BMP7 pathway. The intermediate signaling molecules between BMP7 and ERK remain to be determined.
Summary
Our results and hypothesis are summarized in Fig. 3 Fetal kidney is characterized by upregulation of ERK and p38. JNK is downregulated at this stage. ERK and p38 are necessary for normal kidney development, and their blockade leads to low nephron number with or without reduced kidney size. On the other hand, excessive activity of ERK and p38 results in cystic kidney with interstitial fibrosis. Developmental programmes are proposed to be recapitulated in adult disease state. We reported that ERK and p38 are overexpressed and activated in rat diabetic kidney [25,26]. Renal fibrosis, a final common pathway for end stage renal disease, as well as acute kidney injury also recapitulates embryological programmes [27-29]. Inhibition of ERK or p38 may have potential as a therapeutic strategy. Pharmacological systemic inhibition has several difficulties, however. As mentioned above, administration during the embryonic stage affects the normal kidney development. Also, we attempted to treat diabetic nephropathy by an oral MEK inhibitor PD184352, but it tuned out that PD184352 ameliorated hyperglycemia per se precluding the experiment we had planned [30]. Furthermore, administration of an inhibitor of one MAPK may activate another MAPKs as suggested by our organ culture study of pcy kidney or as shown by others [18,31]. Finally, blockade of ERK or p38 in acute kidney injury may aggravate the course by inhibiting regenerative pro cess. Novel drug-delivery technology targeting specific sites of the kidney at the appropriate timepoints may solve these problems in the future.
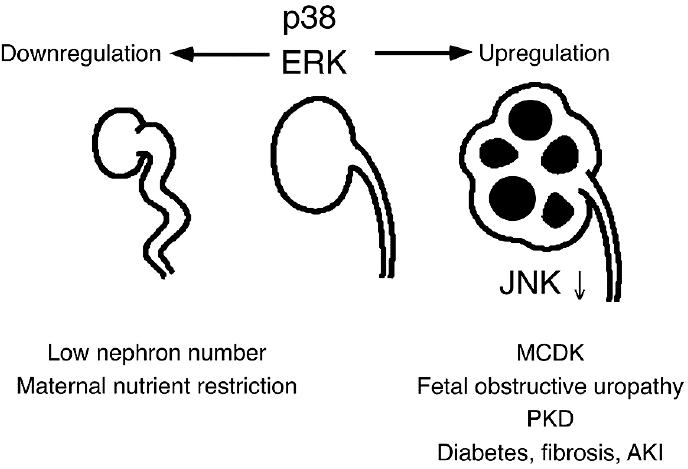
Role of MAPKs in normal and diseased kidneys. p38 and ERK are necessary for normal kidney development. Downregulation of p38 or ERK results in low nephron number with or with- out reduced kidney size as seen in kidneys from the offspring of nutrient restricted dams. Upregulation of p38 and ERK leads to cyst formation and interstitial fibrosis as seen in dysplasia, fetal obstructive nephropathy, and polycystic kidney disease. Adult kidney diseases such as diabetes, fibrosis, and acute kidney injury recapitulate embryological programmes and characterized by fetal pattern of MAPKs expression and activation.
Notes
Conflict of interest
No potential conflict of interest relevant to this article was reported.
Acknowledgements
This study was supported by JSPS KAKENHI grant Numbers 10670757, 17591115, 20591286, and 26461620.